|
|
|
|

| ||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|
Stress and disease tolerance |
|||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|
Breeding for submergence tolerance |
|||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||
Improved varieties that combine high yield with submergence tolerance and elongation ability are needed! |
|
Introduction: The problem!
More than 16 million ha of rice lands of the world in lowland and deep-water rice areas are unfavorably affected by flooding due to complete submergence. The estimated annual economic loss of this is more than US$ 600 Million!
Flood-prone areas in Asia and west Africa
Rice is the only crop plant adapted to aquatic environments because of its well-developed aerenchyma tissues that facilitate oxygen diffusion through continuous air spaces from shoot to root and avoid anoxia development in roots. However, complete submergence due to frequent flooding can adversely affect plant growth and yield.
Two types of flooding cause damages to rice
1. Flash flooding, which results in rapid ascending of water levels with submergence for 1-2 weeks. This type of flooding could also occur after sowing in areas where direct seeding is practiced and could result in substantial reduction in stand establishment.
2. The second type is deepwater and floating rices where water depth exceeds 100 cm and remains at these depths for several months. Plants may become completely submerged for short periods if flooding is severe. Elongation ability of leaves and internodes are essential to keep pace with increasing water levels and escape complete submergence.
Traditional varieties adapted to these environments are low yielding due to their low-tillering ability, long droopy leaves, and susceptibility to lodging, and poor grain quality.
Improved varieties are needed that combine high yield attributes with submergence tolerance and elongation ability. Elongation ability of the coleoptiles of germinating seeds is also considered a desirable trait especially with direct seeding to effect emergence above anaerobic waterlogged soils.
|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|
1. What is the problem of the floodwater environment?
Characterization of floodwater environments in most rice growing areas pointed to gas diffusion as the most important limiting factor under complete submergence. This is because gas diffusion is much slower in water than in air. Oxygen levels in floodwater vary with location and time of day, usually below air saturation during the night but may become supersaturated during the day. Anoxia for 24 h can kill sensitive rice varieties presumably because of the need for O2 for respiration to maintain survival and elongation growth processes.
One important component of survival during submergence is maintenance of carbon assimilation to supply needed energy for maintenance and growth processes. Carbon assimilation is influenced by several factors during submergence, including capacity for underwater photosynthesis, CO2 supply, irradiance and temperature of the floodwater. CO2 supply may be limiting due to both its lower level especially in stagnant water, and the large boundary layer effect. Carbon assimilation of submerged rice plants could also be reduced by low irradiance due to water turbidity or growth of surface algal flocks. Reduction of photosynthesis due to these constraints may impede carbohydrate supply needed for anaerobic or aerobic respiration. |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|
2. Mechanisms of Submergence Tolerance
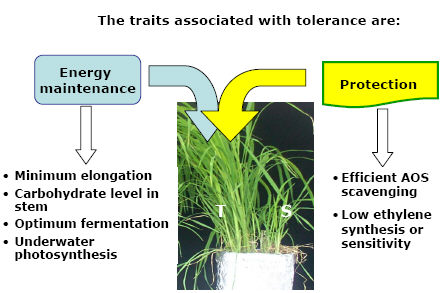
Under flash flooding, few characters were identified as playing a key role in submergence tolerance in rice, the most critical are:
Protective mechanisms as the upregulation of antioxidant system and low synthesis or sensitivity to ethylene during submergence were also found to be useful.
2.1 Stem Carbohydrates Carbohydrate concentration before and during submergence has long been recognized as an important factor in submergence tolerance in rice. For example a strong positive correlation between carbohydrate concentration prior to submergence and tolerance to submergence is commonly observed.
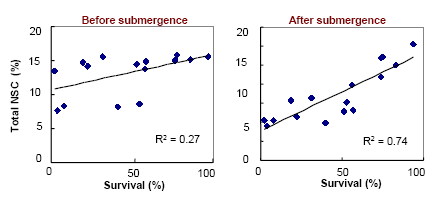
The carbohydrates remaining after submergence is more important than that before submergence
Studies using techniques that alter the concentration of carbohydrates before or during submergence support the significant role carbohydrates plays during submergence e.g. (a) Reduction of light intensity by shading, increasing water turbidity or water depth reduced survival, (b) time of day submergence which affect the diurnal cycle of carbohydrate concentration, (c) CO2 supply which affect underwater photosynthesis (d) seed size; high correlation observed between carbohydrate level and anoxia tolerance of rice seeds (e) seedling age as older seedlings having higher carbohydrate levels and better survival.
Genetic variability in stem carbohydrate content is present among rice cultivars, but greatly influenced by growth conditions before submergence. Selection of genotypes with high stem carbohydrates and with greater ability to conserve it during flooding could provide an increment of submergence tolerance as our recent studies indicated that the level of carbohydrates remaining after submergence is more critical for survival.
2.2 Alcoholic Fermentation (Anaerobic respiration) Alcoholic fermentation (AF) is one of the major metabolic adaptations that plants assume when they are submerged or faced with lack of oxygen. Under anaerobic conditions aerobic respiration shift to a less efficient anaerobic fermentation to provide energy and sustain plant life. Although the amount of ATP (Adenosine triphosphate) produced by this process is very small (5%) compared to ATP produced through aerobic respiration, it is still vital for survival. For rice, this pathway is probably important both under anoxic as well as aerated floodwater where boundary layer limits diffusion of O2 in water and may result in anoxic conditions within tissues. The importance of increased rates of AF during anoxia for plant survival has been demonstrated by several experimental observations.
Examples of these: (1) enzymes of AF often increase under flooding (2) hypoxia pretreatment increased tolerance to anoxia and presumably the induction of AF enzymes (3) Mutants lacking ADH die more quickly under anoxia (4) rates of AF are related to the tolerance of several species to flooding or water logging (5) high sugar supply improved survival, presumably due to continued functioning of AF.
Besides, recent studies with rice plants overexpressing PDC gene (pdc1) further confirmed the importance of AF during submergence. Transgenic lines showed higher activities of PDC enzyme, higher ethanol production and better survival after submergence
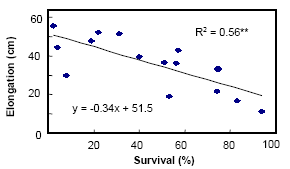
2.3 Stem elongation Under flash flooding stem elongation is not enviable and limited stem elongation growth is found to be associated with cultivars ability to survive flash flooding. This is probably due to energy conservation during flooding for maintenance and survival processes. A strong negative correlation between percent survival and elongation growth is commonly observed.
There is a strong negative correlation between elongation ability and survival
Submergence tolerance of sensitive cultivars was substantially improved when underwater elongation is minimized by application of a gibberellins biosynthesis inhibitor, paclobutrazol. Addition of GA increased elongation and reduced survival of even submergence tolerant lines.
Submergence tolerance and shoot elongation: The 2 samples both on the left and the right show submergence tolerance, the 2 samples in the middle are sensitive to submergence.
2.4. Aerenchyma Formation Aerenchyma comprises gas filled spaces within plant tissue and is considered essential anatomical adaptive trait for survival under flooded conditions. Sufficient substantiation of its role in submergence tolerance was presented by several experiments, in providing a diffusion path of low resistance for the transport of oxygen from shoot to roots in waterlogged soils as well as diffusion of volatile compounds produced in anaerobic soils and plant tissue. In rice the formation of aerenchyma occur both in roots and shoots to provide a continuum from root to shoot
2.5 Role of Ethylene generated during submergence: Submergence-intolerant cultivars usually showed increased levels of leaf chlorosis followed by seedling death during submergence. Ethylene produced during submergence trigger leaf senescence that reduces seedling survival.
This was proved by using an ethylene inhibitor, 1-methyl cyclopropene (MCP). MCP was used to evaluate the effect of ethylene on plant survival, chlorophyll content during and after submergence as well as on enzyme activity and gene expression of chlorophyllase, the first enzyme involved in the chlorophyll degradation pathway. MCP treatment decreased chlorophyll degradation, lowered the activity and gene expression of chlorophyllase enzyme, and improved survival in IR42, a submergence sensitive variety. Retention of chlorophyll by inhibiting ethylene synthesis and action may have improved underwater photosynthesis and provided carbon reserves needed for maintenance metabolism.
Manipulation of ethylene synthesis or sensitivity through molecular approaches may help improve tolerance to flash flooding in rice.
Ethylene gets generated during submergence. Blocking ethylene decreases chlorophyll degradation, increases sugar and starch content and improves survival. (T= tolerant; S= sensitive)
2.6 Post Submergence Events Upon recedes of water level after complete submergence, plants are subjected to both high light intensity and higher oxygen levels. Visual symptoms of injury normally are not evident immediately after desubmergence, but develop progressively during post-submergence.
This post-anoxic injury is caused by generation of reactive oxygen species and toxic oxidative products as acetaldehyde. Oxygen is one possible source of active oxygen species. When O2 gets reduced one electron leaks out from the electron transfer system converting it into superoxide anion (O2-), which in turn produces more active O2 species as hydrogen peroxide (H2O2) and hydroxyl radical (OH.). These highly reactive oxygen species can oxidize unsaturated fatty acids in cellular membranes and intercellular organelles. Recent studies showed that levels of H2O2 and malondialdehyde (a product of lipid peroxidation) were lower in the tolerant cultivar both during and after submergence. This was associated with higher levels of reduced ascorbate antioxidant and activities of enzymes involved in the regeneration of antioxidant metabolites, suggesting the involvement of active oxygen scavenging species in lowering the harmful effects associated with oxygen reentry. Moreover, high light intensity during recovery aggravated the damage due to submergence especially in the intolerant lines.
|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
Symptoms of injury (yellowing of leaves) develop progressively after submergence. This is caused by the generation of active oxygen species (AOS) and toxic oxidative products. |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|
3. Genetics of submergence tolerance Research results suggested both simple and quantitative inheritance for submergence tolerance. In one study, the involvement of at least three genes was reported with low to moderated broad sense heritability.
In another study, analysis of segregating F2 and backcross populations made between tolerant and intolerant lines suggested the involvement of one major dominant gene in the submergence tolerance of three tolerant lines, FR13A, BKNFR (76106-16-0-1-0) and Kurkaruppan. In a set of double haploid population developed between a sensitivee and a tolerant line and screened in Thailand under high irradiance in the field, a strong bimodal distribution was observed with equal number of individuals in the tolerant and intolerant categories suggesting the involvement of a single gene in submergence tolerance and agreed with the model discussed above. However, when the same population was screened under low irradiance in Philippines, results suggest multiple genes or a more complex inheritance. These differences in response may be due to differences in environmental conditions. When plants are grown at high irradiance carbohydrate levels would have been high and possibly eliminating its effect as a limiting factor. |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|
4. Mapping of QTLs associated with submergence tolerance: Using a cross between an indica submergence tolerant line (IR40931-26) and a sensitive japonica line (PI543851), a major QTL associated with submergence tolerance, designated Sub1, was mapped to chromosome 9. This QTL accounted for 70% of the phenotypic variance of submergence tolerance in the population studied, which is extremely high for a QTL and again suggests the involvement of a major gene. The donor line for this QTL was derived from the world most submergence tolerant line, FR13A and with similar level of tolerance.
Sub1 locus was recently fine mapped and sequenced and the genes responsible for tolerance were identified as a cluster of Ethylene Responsive Elements (ERFs). Two of these genes, Sub1B and Sub1C, are always present in the Sub1 region of all rice accessions analyzed so far, but the presence of Sub1A is variable with two alleles; a tolerance-specific allele ‘Sub1A-1’ and an intolerance-specific allele ‘Sub1A-2’. Overexpression of Sub1A-1 in an intolerant cultivar conferred enhanced tolerance, indicating that Sub1A-1 is a primary determinant of submergence tolerance. DNA markers that are specific to Sub1 were developed and used in marker assisted backcrossing (MAB) to incorporate Sub1 into popular, but intolerant varieties. This locus was now introgressed into few widely grown cultivars using MAB such as Swarna, IR64 and Sumba Mahsuri and efforts are on-going to incorporate it into additional “mega” varieties. The new varieties maintain the high yield and other agronomic properties of the recurrent parents and are tolerant to submergence. Cultivation of these varieties is expected to provide protection against damaging floods and increase the food security for farmers in flood-prone areas.
Four minor QTLs were also identified on other chromosomesand are currently being pursued with the objective of pyramiding them with Sub1 for higher levels of tolerance. Improving our understanding of the physiological and molecular aspects of tolerance mechanisms will help in identification of additional candidate genes involved to be used as markers for breeding. |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|
6. How to screen for submergence tolerance and avoidance:
Tolerance to complete submergence: Duration of submergence depends on several factors the most important of which are seedling age, water temperature, turbidity and light intensity. At IRRI, screening is commonly performed 30 d (= days) after transplanting of 21 d old seedlings or 30 d old seedlings with direct seeding. Both are conducted in artificial ponds. Water level should be raised fast and maintained to a depth of at least 1 m for 10 to 14 d as the standard period of treatment. To avoid severe conditions e.g. when water temperature is high or water is turbid, it is better to monitor the performance of the sensitive check and drain the water once the base of its stem start to soften.
Submergence avoidance (Elongation ability) Treatment should start 30 d after transplanting of 21d old seedlings. Water level is usually increased at a rate of 5 cm/day up to 100 cm. The evaluation can be conducted at any time while the water level is rising or after reaching the maximum height of 100 cm. Lines that continue to elongate and keep pace with the rising water could be selected and evaluated for other agronomic and quality traits.
Submerged field at IRRI
|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|
7. Breeding for submergence tolerance
7.1 Problems in genetic improvement of flood-prone rice Excess water is common in many rice production areas, ranging from rainfed lowland to the very deeply flooded areas of the large river basins with uncontrolled floods where deepwater or floating rice is grown. Conditions vary from temporary submergence of 1-10 days, which affect both rainfed lowlands and deepwater rice; long periods of standing water; or daily tidal fluctuations that sometimes cause complete submergence. Crops grown in these areas are also affected by adverse soil conditions such as acidity, salinity, minimal toxicities, and low soil fertility.
In deepwater (80-100 cm) and floating-rice (>100 cm) areas, soil and water management is difficult. For deepwater areas where the land is fertile, it could be feasible to double the yield with improved varieties. Genetic improvement of floating rice is difficult because of the high energy cost of fast elongation and may not be a current priority at IRRI.
In submergence-prone areas (10-12 days under water) with no major soil problems, yield can also be doubled with the use of submergence tolerant improved varieties. These areas have good yield potentials because of the fertile soil and minimal occurrence of pests and diseases.
7.2 Target traits for breeding
I. Common traits needed in both elongating and submergence tolerant lines:
II. Elongating rice for medium-deep water
III. Elongation
IV. Submergence
V. Submergence avoidance
* = Screened for at IRRI, ** = cannot be done at IRRI, *** = possible at IRRI but not done so far
7.3 Donors for tolerance to excess water stress and other related traits
Submergence tolerance FR13A, Kurkaruppan and Goda Heenati are the major donors used for submergence tolerance in breeding program at IRRI. Some studies suggested that Goda Heenati have different genes for submergence tolerance, which warrant further investigation. However, it might be useful to utilize them simultaneously to conserve genetic diversity. Other lines identified as tolerant to flash flooding: FR43B, Thalavu and BKNFR76106-16-0-1-0. More recently we have found that IR49830-7-1-2-1-3 is an excellent donor with high yielding ability.
Submergence avoidance (Elongation ability) Many traditional varieties for deep water rice and floating rice have been used as donor from various countries. Even some lines of wild rice have been also utilized as donors of elongation ability. Although their genes for elongation ability have not been examined carefully, those donors might be useful in broadening the genetic background. Examples: Jalmagna, Baisbish and Rayada 16-3.
Other target traits Most other useful phenotypic traits are available in improved irrigated rice varieties, such as leaf traits of NPT (New Plant Type), tolerance to diseases, less tillering, etc. Wild relatives of rice could also be used to transfer some useful traits such as the elongation ability of O. rufipogon that has been explored to develop deep water lines. Mutation breeding could be explored to improving the grain quality and plant type of the existing landraces.
7.4. Breeding of rice varieties for excess-water condition and future prospects Based on previous studies on genetics of elongation ability and submergence tolerance, the number of genes involved seem to be relatively small (1 major and 4 minor genes in either case), however, breeding for improved rice varieties with submergence tolerance or elongation ability has been rather slow. This may be partially due to the complexity of the environment and the existence of other site-specific stresses that are commonly not accounted for during screening. Artificial screening ponds and rapid generation advancement are necessary to accelerate breeding efficiency. IRRI acquired some of the needed facilities that could be shared with NARES.
IRRI and NARS have been working on developing germplasm for elongation ability and submergence tolerance. Some of the lines developed have the same level of either elongation ability or submergence tolerance as the donor lines and with better yield and plant type. For example, the new plant types with facultative elongation ability, broad and thick leaves, heavy panicles and stiff culms for deep-water conditions has been developed at IRRI and are available upon request or through INGER. Though, superior varieties have been developed, dissemination is not always successful. The breeders’ expectations of an ideal plant type are not always the same as farmers’. Breeder must understand and analyze the current and future needs in target areas before designing a specific ideotype.
The following are some examples where this proves to be important: In Thailand, for instance, it took 20 years to release the superior variety, Prachinburi2”. The Rice Research Institute of Thailand produced two new breeding lines, HTAFR81042-4B-7-1 and HTAFR84038-B-5-0-1. HTAFR81042-4B-7-1 was named as “Prachinburi2” which was recently released. This variety has a yield of 3.5 t ha-1 at 100 cm water depth and about 5 t ha-1 under irrigated conditions in farmer’s fields. Its wide acceptance by farmers could be attributed to its high yield advantage as well as good quality for processing of rice noodles.
In Indonesia, some breeding lines tolerant to low soil pH were developed. They yield about 4 t ha-1 but farmers in Central and South Kalimantan did not like the size and shape of its grains. Breeders developed varieties with the same level of tolerance to low pH but with slender and fine grain shape and lower yield of only 3t ha-1. Farmers adopt this variety despite the sacrifice in yield of up to 1t ha-1. |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|
Click here to see a list of some germplasm developed at IRRI for flash flooding tolerance: grain yield during 1999 and 2001. |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|
|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||
Let's conclude |
|
Summary
• Floodwater environments have been characterized. Limited gas diffusion, light penetration and temperature and turbidity of the flood water are identified as important factors that affect survival
• Biology and genetics of tolerances to flash-flooding are fairly well understood and this knowledge will facilitate designing suitable screening criteria and breeding strategies.
• The major physiological traits involved in submergence tolerance are maintenance of high carbohydrate concentration, minimum elongation growth, optimum rates of alcoholic fermentation, upregulation of antioxidant system when water recedes and low synthesis or sensitivity to ethylene during submergence.
• A major locus for submergence tolerance (Sub1) is tagged and is being used in marker assisted breeding (MAS). New varieties with Sub1 showed similar agronomic and quality traits as their parents but substantially higher yields when submerged. The Sub1 locus could possibly be intorgressed into any popular variety within 2-3 years using MAB.
• Donor lines with better agronomic traits and new breeding tools are now available. Useful alleles of candidate genes could be tagged and used for MAS breeding. These genes/alleles need to be incorporated into varieties that are most popular in the target environments to accelerate adoption and diffusion.
• Beside tolerance to flash flooding, other traits are also essential for high and stable productivity in flood-prone areas. These include tolerance to flooding during germination, tolerance to stagnant or water logged conditions and higher regeneration ability after water recedes. Efforts are now ongoing to develop cultivars with enhanced tolerance to these conditions
|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|
Additional information: Deepwater breeding
Click on the icon below to open a PDF file with information on Deepwater Rice (by S. Yanagihara)
PDF Deepwater rice |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||
Next lesson |
|
In the next lesson, you will learn how to screen for drought tolerant varieties. |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|
|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||


.gif)